Research Article
Isolation, Culture, Identification and Differentiation of Canine Amniotic Mesenchymal Stem Cells in Vitro 
2 Inner Mongolia Maternal and Child Health Hospital, Huhhot, China
 Author
Author  Correspondence author
Correspondence author
Animal Molecular Breeding, 2017, Vol. 7, No. 1 doi: 10.5376/amb.2017.07.0001
Received: 20 Apr., 2017 Accepted: 25 Apr., 2017 Published: 28 Apr., 2017
Zhang Y.K., Bai L.H., Li Y., Liu M.N., Gao G.F., and Zhi D.F., 2017, Isolation, culture, identification and differentiation of canine amniotic mesenchymal stem cells in vitro, Animal Molecular Breeding, Vol.7, No.1, 1-13 (doi: 10.5376/amb.2017.07.0001)
Recent years, people are paying more and more attention to amniotic membrane tissue which has a rich resource and no ethical restrictions. Canine amniotic mesenchymal stem cells (cAMSCs) were collected by enzyme digestion method from canine amniotic membrane tissues. The cell morphology was continuously observed at different passages and cell growth curves of passage 3 and passage 9 were drawed. Stem cell related proteins and genes were identified by immunofluorescence and RT-PCR. The multiple differentiation potential of cAMSCs was identified by osteogenesis and adipogenesis. This study showed that the cAMSCs was long fusiform and could be continuously subcultured in vitro. The immunofluorescence results showed that Vimentin and SSEA-4 were positively expressed. The RT-PCR results showed that stem cell related genes OCT4, SOX2 and NANOG were positively expressed too. The cAMSCs could be induced to osteoblasts and adipocytes in vitro. The markers associated with osteogenesis and adipogenesis, such as COL1A1 and LPL, were positively expressed after differentiation. The results showed that the cAMSCs could be successfully isolated and cultured in vitro. Stem cell related proteins and genes were positively expressed. And the cAMSCs has the multiple differentiation potential. This study showed that cAMSCs could be a rich source of stem cells in veterinary. Furthermore, cAMSCs may be useful as a cell therapy application for veterinary regenerative medicine.
Introduction
Mesenchymal stem cells (MSCs) are multipotent stem cells originating from extraembryonic mesoderm (Fortier 2005; Dominici et al., 2006). With self-renewal and multiple differentiation capability, MSCs under different induction conditions can differentiated into kinds of somatic cell types including osteoblast, neurocyte and pancreatic cells (Csaki et al., 2007; Vieira et al., 2010; Guercio et al., 2012). MSCs were originally obtained from bone marrow (Kortesidis et al., 2005). In recent years, a large number of studies have shown that MSCs can be successfully isolated from muscle, adipose, umbilical cord blood, amniotic fluid and placenta (Kisiel et al., 2012; Takemitsu et al., 2012; Ryu et al., 2012). Therefore, MSCs have the potential to provide the field of regenerative medicine and tissue engineering a particularly attractive tool for cell-based therapies.
The placenta plays an important role during pregnancy in the nutrient uptake, waste elimination, and gas exchange during pregnancy. The fetal membrane is a part of the placenta which surround and protect a developing fetus. The typical structure of the placenta is a three-layer membrane including the deciduas, chorion and amnion without blood vessel and lymph vessel (Parolini et al., 2008; Bárcena et al., 2011). Canine amnion tissues are a rich source of MSCs with easy acquisition. Generally, placenta tissues are often discarded as a kind of medical waste following birth (Serikov et al., 2009; Murphy et al., 2010). So, canine amnion tissues are readily available without any invasive or ruthless methods. This means that MSCs derived from amnion tissues have minimal ethical and legal constraints.
The safety and efficacy of MSCs-based therapy for humans must be evaluated in preclinical animal models (Csaki et al., 2007). Dogs has been used in the evaluation of new therapeutic drugs or novel treatments (Schneider et al., 2008; Naggara et al., 2010). The similarities in regard to anatomical and physiological characteristics make dogs a very attractive animal model (Little and Smith 2008; Chu et al., 2010). Therefore, dogs have been gaining wide attention as a new superior laboratory animal model for the research of human diseases. Recently, canine MSCs have been used as an alternative treatment for canine regenerative medical applications, such as bone problems and osteoarthritis (Crovace et al., 2008; Guercio et al. 2012).
In the present study, cAMSCs were isolated from the canine amnion in vitro. We evaluated the biological characteristics, stemness protein and gene expression, immunophenotype and multiple differentiation potential of cAMSCs. Based on our findings, the self-renewal and highly multiple differentiation capacity of cAMSCs suggest potential clinical applications in canine regenerative medicine studies.
1 Materials and Methods
1.1 Animals
All animal procedures were approved by the Ethics Committee of Inner Mongolia Agricultural University. Four healthy female adult mixed-breed dogs (aged 2 to 4 years) were used in this study. Canine placentas were obtained through caesarean section in Inner Mongolia Agricultural University Veterinary Hospital. In the cesarean section, anesthesia was induced with propofol through intravenous injection and maintained with low-flow inhalation of isoflurane. All animal tissues were only used in this study for isolating stem cells from amniotic membrane.
1.2 cAMSCs isolation and culture
cAMSCs were isolated as previously described with slight modification (Tsuji et al., 2010; Diaz-Prado et al., 2010). Canine placentas were obtained through caesarean section from healthy female canines. Briefly, the amniotic membrane (AM) was separated from the chorionic membrane mechanically. Under aseptic condition, the collected AM were rinsed several times with phosphate-buffered saline (PBS; Hyclone) containing 100 U/ml penicillin and 100 g/ml streptomycin (Gibco). The washed AM cut into small pieces with ophthalmic scissors. AM was composed of a simple epithelium layer and an avascular stroma layer. Firstly, in order to get rid of epithelial cells, the minced AM was digested with 0.05% trypsin-EDTA at 37°C for 60 minutes. After digestion, the AM was washed with PBS for 3 times. Secondly, the AM was then subjected to a second enzymatic digestion in Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F12; Gibco) containing 1 mg/ml collagenase type I (Gibco) at 37°C for 60 minutes. The cells were collected by filtration of 70 μm cell strainer and centrifugation at 350×g for 5 min. The harvested cell pellet was resuspended in the complete medium composed of DMEM/F12 supplemented with 10% fetal bovine serum (FBS; Hyclone) and 100 U/ml penicillin and 100 g/ml streptomycin. The cells were seeded into100 mm polystyrene cell culture dish (Corning) and incubated in a humidified atmosphere with 5% CO2 at 37°C. The nonadherent cells were removed after 72 h of culture and the adherent cells passaged as 1:3 when reaching 80-90% confluency. The culture medium was changed 3 times a week.
1.3 Identification of cell surface markers by flow cytometry
Take the third generation of cells, first with 0.25% trypsin digestion, then DPBS resuspended, The cell density was 1 × 107 cells / mL; pipette 100 μL of cell suspension in each flow cytometry tube, 20μL of PE-labeled CD29, CD34, CD49d, and CD73 were added to each tube and FITC-labeled CD45, HLA-DR monoclonal antibodies, incubate for 30 min at room temperature in the dark, then added 2 mL of DPBS containing 0.09% NaN3, mixing, centrifuge at 1500 rpm for 5 min, then the supernatants were removed; finally, weight drop with 500 μL DPBS, flow cytometry detection, 10,000 cells were collected per sample,results were analyzed using Cell Quest software.
1.4 Growth curves and population doubling time
The 3rd, 6th, and 9th passages of cAMSCs were harvested using 0.25% trypsin-EDTA when reaching 90% confluence and then seeded into 24-well plates at 3×104 cells/well. The cells of each group were randomly sampled for digestion and counted at the same time point every day. The growth curves were established, and population doubling time (PDT) was calculated using the formula t=lg2/(lgNt−lgN0), where Nt represent the final harvest cell number, N0 represent the initial seeding cell number.
1.5 Immunostaining
Immunofluorescence was used to identify the cAMSCs markers Vimentin and SSEA-4. When the 3th passage cells reaching 40-50% confluence, the culture medium was removed. The cells were washed twice with PBS and fixed with 4% paraformaldehyde overnight at 4°C, washed several times with PBS. Then the cells were permeablized at room temperature for 2 h in 0.1% Triton X-100 diluted in PBS and washed three times with PBS. The cells were blocked with normal goat serum for 2 h at 37°C. Primary antibodies Vimentin and SSEA-4 (1:100) were added and incubated overnight at 4°C. After washing three times with PBS, the cells were incubated with FITC labeled secondary antibodies IgG (1:20) for 30 min at room temperature in dark, and cells were washed three times with PBS. Finally, DAPI solution diluted 1:100 in PBS was added to the wells, then the cells were incubated at room temperature for 15 min. The cells were washed three times with PBS to remove unbound DAPI. Images were captured with a confocal microscope.
1.6 Reverse transcriptase-polymerase chain reaction (RT-PCR)
Expressions of specific genes for differentiation and stemness genes were determined by RT-PCR as reported earlier (Li et al., 2014). Briefly, total RNA from different samples was isolated using RNAiso Plus (TaKaRa, Otsu, Japan, Cat. #9108). RNA concentration was quantified by absorbance at 260 nm using a spectrophotometer. The first strand complementary DNA (cDNA) was synthesized by 1 mg of total RNA for reverse transcription using PrimeScript RT reagent Kit with gDNA Eraser (Perfect real time) (TaKaRa, Dalian, China). Reaction for reverse transcription was proceeded according to manufacturer’s protocol with the following procedures: 42°C for 2 min (gDNA remove); 37°C for 15 min (reversely transcribe); 85°C for 5 sec (inactive the reverse transcriptase). The PCR primers and the reaction conditions used are shown in Table 1. GAPDH was used as housekeeping marker genes at all stages. Reactions without template cDNA were used as a control for PCR. Thermocycling was carried out in a final volume of 20 μl containing 2.0 μl cDNA, 10 μM forward and reverse primers each, and 10 μl of Taq DNA polymerase (TaKaRa, Dalian, China). The PCR primers and the reaction conditions used are shown in Table 1. GAPDH was used as housekeeping marker genes at all stages. Reactions without template cDNA were used as a control for PCR. The PCR products were separated on a 2% agarose gel with ethidium bromide staining.
.png) Table 1 List of PCR primers |
1.7 In vitro differentiation potential
When the 3th passage cells reaching 80% confluence, the culture medium was changed to the specific differentiation medium. Osteogenic differentiation medium contained LG-DMEM supplemented with 10% FBS, 10 nM dexamethasone, 10 mM β-glycerophosphate, 50 mM ascorbic acid 2-phosphate (Sigma, USA). After 4 weeks, the differentiation of cells was assessed morphologically and detected by staining with Alizarin Red S and Alkaline Phosphatase Staining, which indicate the calcium deposition and marker enzyme of mature osteoblast. Adipogenic differentiation medium contained LG-DMEM supplemented with 10% FBS, 1 μM dexamethasone, 10 mg/L insulin, 500 μM 3-isobutyl-1-metyl-xanthine, 100 μM indomethacin (Sigma, USA). After 3 weeks, the differentiation of cells was assessed morphologically and detected by staining with Oil Red O, which indicate the presence of lipid droplets in cells. The differentiation medium was changed 3 times a week. Basal culture medium was used in each group as a control condition.
1.8 Alizarin red S staining
Differentiation medium was aspirated from each well. The cells were washed with PBS several times and fixed with 70% ethanol for 1 h at 4°C, washed several times with distilled water. The cells were stained with 0.1% Alizarin Red S (Sigma, USA) diluted in distilled water (w/v) and then incubated at room temperature for 20 min. And the cells were washed 3 times thoroughly with distilled water. The cells were observed and imaged with phase contrast microscope.
1.9 Alkaline phosphatase staining
ALP staining was performed using cell Alkaline Phosphatase (cALP) stain kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The cells were washed twice with PBS and fixed with 4% paraformaldehyde for 1 h at 4°C, washed several times with distilled water. The cells were incubated with cALP stain kit in dark at room temperature for 20 min. After rinsing with distilled water, the cells were observed and imaged with phase contrast microscope.
1.10 Oil red O staining
The cells were washed with PBS several times and fixed with 4% paraformaldehyde for 1 h at 4°C, washed several times with distilled water. The cells were incubated with 0.16% filtered Oil Red O (Sigma, USA) diluted in isopropanol (w/v) at room temperature for 10 min. The stain solution was aspirated and stains in the wells were solubilized with100% isopropanol. The cells were observed and imaged immediately after staining under phase contrast microscope.
2 Results
2.1 Isolation and culture
Canine placentas were obtained through caesarean section. The AM was mechanically separated from the chorionic membrane and moved to a sterile culture dish (Figure 1A). Then the primary cAMSCs were isolated and cultured from the collected AM tissues. The speed of cell attachment was fast. A few discrete cell colonies started emerging and attached to the plastic culture dish at 24 h after initial seeding, but most cells were still suspended in the medium (Figure 1B). The culture medium was first changed at 72 h to remove the nonadherent cells. Most cells had attached to the culture dish with various shapes, which were spindle-shaped, round or polygonal. The proliferation rate was fast and the cells reached 90% confluency after 6 d of seeding. (Figure 1C). The 3th passage cells displayed a typical spindle-shaped and fibroblast-like morphology with radiating and whirlpool-like migrating patterns, which is similar to bone marrow mesenchymal stem cells. The density, size and purity of the 3th passage cells increased significantly after subculture. (Figure 1D). The third passage cells were stored in liquid nitrogen for future use.
.png) Figure 1 Primary culture of canine amniotic mesenchymal stem cells Note: A: Canine amniotic membrane tissue obtained through caesarean section; B: Cells started emerging and attached to the plastic culture dish at 24 h after initial seeding (×100); C: Cells attached to the culture dish with spindle-shaped, round or polygonal shapes. (×100); D: The 3th confluent passage cells in primary culture displayed a typical spindle-shaped and fibroblast-like morphology with radiating and whirlpool-like migrating patterns (×100) |
2.2 Growth curve
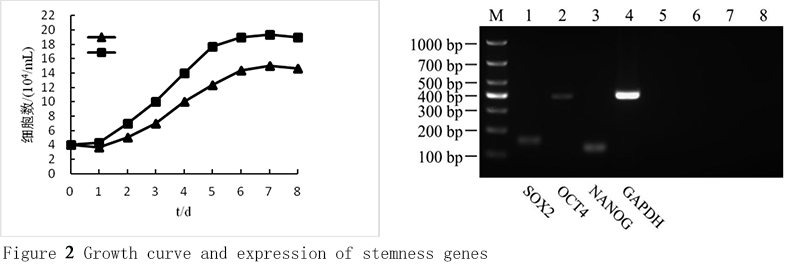
To determine self-renewal capacity of cAMSCs, the cells at 3th, 6th and 9th passages were separately cultured in 24-well plates at 3×104 cells/well, and counted at the same time point every day. The cell growth curves were drawn based on the numbers of the cells. The results demonstrate that the growth curves of 3th, 6th and 9th passage cells share a similar typical S-shape (Figure 2A). The cell growth was slow at 1-2 d, which was called incubation period. At logarithmic growth phase of 3-6 d, the cells grow quickly. The cell growth was slowed down after entering the stationary phase of 7-8 d. Compared with 6th and 9th passage, the proliferation rate of 3th passage cells was strong. The 9th cells changed their morphology with retarded proliferation rate and increased size.
|
Figure 2 Growth curve and expression of stemness genes Note: A: Growth curve of cAMSCs at 3th, 6th and 9th passages; B: Expression of stemness genes by RT-PCR analysis. Lane 1 DL1000 marker, lane 2 SOX2 (152 bp), lane 3 OCT4 (388 bp), lane 4 NANOG (127 bp), lane 5 GAPDH (392 bp), lane 6 SOX2 negative control, lane 7 OCT4 negative control, lane 8 NANOG negative control, lane 9 GAPDH negative control. |
2.3 Expression of cell surface markers in cAMSCs
The results of flow cytometry showed that, third-generation cAMSCs had high expression CD29、CD49d and CD73, respectively 99.85%、87.61% and 98.79%(Figure 3D; Figure 3E; Figure 3F), low expression CD34, CD45 and HLA-DR, respectively 0.61%, 0.24% and 0.29% (Figure 3A; Figure 3B; Figure 3C), Figure 2G is a blank control group. The results showed that the purity of cAMSCs was high.
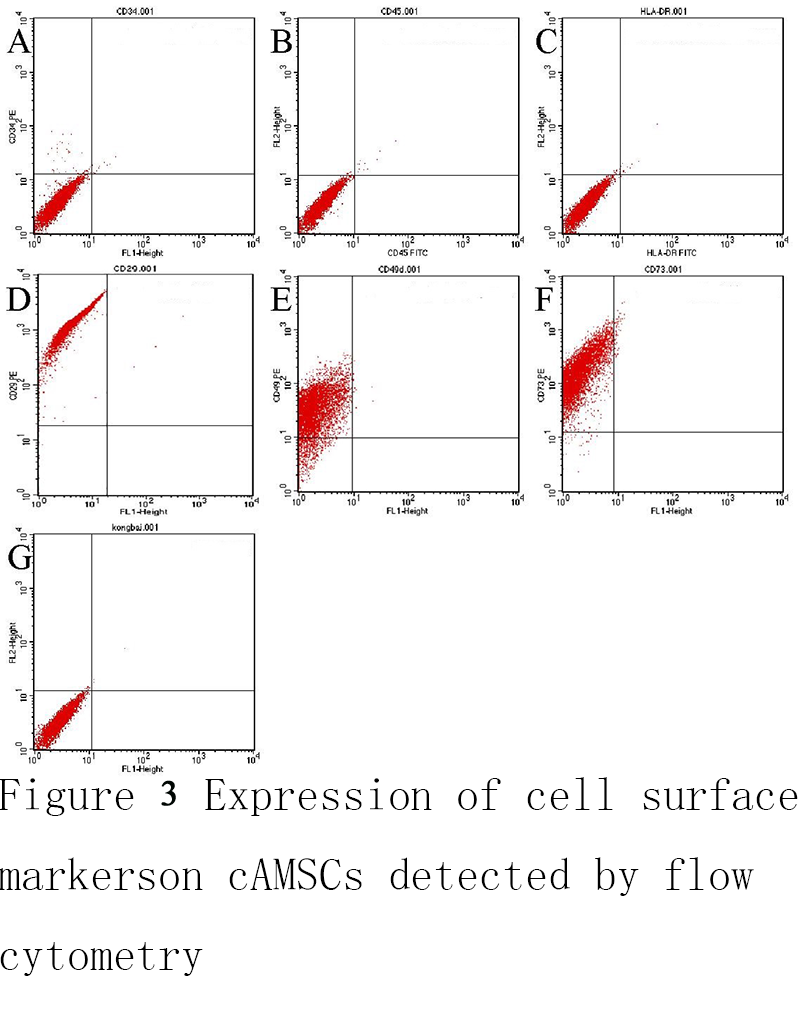 Figure 3 Expression of cell surface markers on cAMSCs detected by flow cytometry |
2.4 Expression of stemness genes
Expressions of SOX2, OCT4, and NANOG were determined by RT-PCR analysis to characterize the stemness genes of cAMSCs. Total RNA was isolated from 3th passages cells. The RT-PCR showed that the cells expressed the transcription factors of these genes. The PCR products of all these genes were sequenced and the amplimer sequences were aligned and analyzed using the Basic Local Alignment Search Tool. The sequence of SOX2 had 100% identity with Canis lupus familiaris SOX2 mRNA. Alignment of OCT4 sequence showed 100% homology with Canis lupus familiaris homeobox transcription factor OCT4 mRNA. NANOG blast showed 100% homology with Canis lupus familiaris Sox2 mRNA (Figure 2B).
2.5 The expression of vimentin and SSEA-4 was detected by immunofluorescence
The third generation of cAMSCs were adhered to the glass slide, were tested for immunofluorescence. The results showed that the nucleis were blue fluorescence by DAPI staining, and the cytoplasms were green fluorescence, which indicated that Vimentin and SSEA-4 were positive expression in the cytoplasm of cAMSCs, the control group with DPBS instead of the primary antibody was negative expression (Figure 4).
.png) Figure 4 Expression of Vimentin and SSEA-4 on cAMSCs detected by immunofluorescence Note: (A)(E) The cytoplasm was green fluorescent, indicating that Vimentin and SSEA-4 were positive in cytoplasm of cAMSCs (100×) (B)(F) The nuclei stained with DAPI was blue fluorescence (100×); (C)(G) Cell membrane and cytoplasm of the fluorescence generated by the composite image(100×); (D)(H) The control group with DPBS instead of primary antibody was negative (100×) |
2.6 Induction of osteogenic differentiation
To investigate the osteogenic differentiation potential of cAMSCs, the cells were cultured in osteogenic differentiation medium for 4 weeks. The cells began to alter morphology after 10 d of incubation until last day of induction differentiation (Figure 5A). On 28 d of culture, Alizarin Red S and Alkaline Phosphatase Staining were used to assess the osteogenic differentiation. Under differentiation conditions, Alizarin Red S staining was positive under osteogenic conditions, which confirmed the existence of calcium deposits. Meanwhile, there was strong and positive Alkaline Phosphatase Staining. However, under both control conditions, negative staining was observed under Alizarin Red S and Alkaline Phosphatase Staining. The expressions of specific genes associated with osteogenesis, such as SPARC and COL1A1, were determined by RT-PCR analysis. The result of RT-PCR shows that these genes were strongly expressed under differentiation conditions compared to controls.
.png) Figure 5 Osteogenic differentiation of cAMSCs Note: Alizarin Red S and Alkaline Phosphatase Staining were used to assess the osteogenic differentiation after 4 weeks’ culture. In Figure 5, (A) Cells were cultured in osteogenic differentiation medium and Alizarin Red S staining was positive under differentiation conditions (×100); (B) Negative staining with Alizarin Red S was observed under control condition (×100); (C) Alkaline Phosphatase Staining was positive under differentiation conditions after 28 d culture in osteogenic differentiation medium (×200); (D) Alkaline Phosphatase Staining was negative under control condition (×200); (E) Expression of COL1A1 by RT-PCR analysis. Lane 1 DL1000 marker, lane 2 GAPDH undifferentiation (392 bp), lane 3 COL1A1 undifferentiation (124 bp), lane 4 GAPDH differentiation (392 bp), lane 5 COL1A1 differentiation (124 bp), lane 6 GAPDH negative control, lane 7 COL1A1 negative control |
2.7 Induction of adipogenic differentiation
To investigate the adipogenic differentiation potential of cAMSCs, the cells were cultured in adipogenic differentiation medium for 3 weeks. The cells began to alter morphology after 10 d of incubation until last day of induction differentiation (Figure 6A). On 21 d of culture, Oil Red O Staining was used to assess the adipogenic differentiation. Under differentiation conditions, Oil Red O staining was positive under adipogenic conditions, which confirmed the existence of lipid droplets in cells. However, under control condition, negative staining was observed under Oil Red O Staining. The expressions of specific genes associated with adipogenesis, such as LPL and Leptin, were determined by RT-PCR analysis. The result of RT-PCR shows that these genes were strongly expressed under differentiation conditions compared to controls.
.png) Figure 6 Adipogenic differentiation of cAMSCs Note: Oil Red O staining was used to assess the adipogenic differentiation after 4 weeks’ culture. (A) Cells were cultured in osteogenic differentiation medium and Oil Red O staining was positive under differentiation conditions; (B) Negative staining with Oil Red O was observed under control condition; (C) Expression of LPL by RT-PCR analysis. Lane 1 DL1000 marker, lane 2 GAPDH undifferentiation 392 (bp), lane 3 LPL undifferentiation 134 (bp), lane 4 GAPDH differentiation 392 (bp), lane 5 LPL differentiation (134 bp), lane 6 GAPDH negative control, lane 7 LPL negative control; (D) Expression of LEPTIN by RT-PCR analysis. Lane 1 DL1000 marker, lane 2 GAPDH undifferentiation (392 bp), lane 3 LEPTIN undifferentiation 101 bp), lane 4 GAPDH differentiation (92 bp), lane 5 LEPTIN differentiation (101 bp), lane 6 GAPDH negative control, lane 7 LEPTIN negative control.
|
3 Discussion
3.1 Isolation and culture of cAMSCs
The structure of placenta consists of three layers including amnion, chorion, and decidua. The inner layer of placenta is amnion, a thin avascular membrane, which are comprised of an epithelial monolayer and an avascular stroma (Parolini et al., 2008; Bárcena et al., 2011). The canine amnion is a highly abundant, inexpensive and readily available tissue that is often discarded as a kind of medical waste post-partum. There are some reports regarding the isolation and characterization of MSCs from human placenta tissues. Under different induction conditions, these cells have the capacity to differentiate into kinds of somatic cell types which are originating from all three germ layers, including endoderm, mesoderm and ectoderm (Csaki et al., 2007; Vieira et al., 2010; Guercio et al., 2012). The MSCs from placenta tissues have many advantages in cell therapy, such as low immunogenicity (Lim et al., 2007), anti-inflammatory properties, and minimal ethical and legal constraints. That makes them to be considered as a preferable alternative source of MSCs in the field of regenerative medicine. Dogs has been used as an animal model in the evaluation of new therapeutic drugs or novel treatments for several human diseases. In veterinary medicine, the application of MSCs has rapidly emerged cell-based therapies. Bone marrow-derived and adipose tissue-derived MSCs are most frequently used in experimental and clinical studies, since these kinds of MSCs have been best characterized. Therefore, new source of MSCs needs to be established.
The structure of amnion consists of five layers including epithelial cells monolayer layer, basement membrane, compact layer, fibroblast layer and spongy layer. So, when the separation AMSCs, need to use the pancreatin enzyme digestion to remove epithelial cells on the surface of amnion, the use of collagenase digestion of the rest of the amnion, cell suspensions obtained by adherent culture obtained AMSCs. But due to the complexity of the organizational structure of the placenta, the degree of enzyme digestion can not be precisely controlled, primary cultured cells may be mixed with other types of cells. These cells proliferation ability is weak, they take a long time when the cell passage of trypsin digestion, therefore, cAMSCs can be further purified by cell passage. Another method is screening of cAMSCs using flow cytometry, using cell surface markers were screened, however, existing studies have not found available for single or untied surface markers, therefore unable to guarantee the resulting that cell sorting purity of the cells and cell sorting method for simplicity.
In this study separated cAMSCs method with hAMSCs in the same way, first, digestion of epithelial cells on the amniotic surface was performed using trypsin, second, by collagenase typeⅠdigestion amniotic organization makes it the individual cells. Most of the cells were suspended in the culture medium after 24 hours, but we can be observed a few morphologically irregular adherent cells, culture medium was changed 72 hours later and the unattached cells were removed, a small number of cells were ovoid, polygonal, most of the cells for long spindle. The cells were purified after twice cell passage, most of the third-generation cells showing long fusiform, single shapes, submitted typical fibroblast whirlpool or flamboyancy growth.
3.2 Biological characteristics of cAMSCs
At present, canine amniotic mesenchymal stem cells have no uniform identification standard, the literatures have been identified by reference to the standard of human amniotic mesenchymal stem cells. In 2006, International Society for Cellular Therapy rule hMSCs three minimum appraisal standard: (1) Can be attached to plastic culture plate growth (2) The surface markers of cMSCs analysis indicated the positive expression of CD73, CD90, CD105, and no expression of CD11b, CD14, CD19, CD29a, CD34, CD45 and HLA-DR (3) Can be inducing include osteogenic differentiation, adipogenic differentiation, chondrogenic differentiation (Dominici et al., 2006). The same reference to the above criteria, by immunofluorescence, flow cytometry and RT-PCR, the immunophenotype and gene expression of cAMSCs were identified, and the differentiation ability of cAMSCs into three kinds of germ layer cells was explored.
Vimentin is mainly expressed in stromal cells, is the backbone of its cytoskeleton of a major intermediate filament fibers, present in the endothelial cells, fibroblasts and leukocytes and other mesodermal-derived cells (Coulombe et al., 2004); stage specific embryonic antigens (SSEA-4) is a glycoprotein on the surface of early embryonic cells, expressed only in undifferentiated early embryonic cells, and is an important marker to identify cell pluripotency. In this study, by immunofluorescence technology proved Vimentin and SSEA-4 in cAMSCs was positive expression, indicating that cAMSCs with both stromal cells and embryonic stem cells’ characteristics.
Recent research demonstrates, by flow cytometry, cAMSCs had high expression CD29, CD44, CD73, CD90, CD105 and MHC-I, low expression CD34, CD45, CD14, CD117, CD133 and MHC-II, meantime cAMSCs had expression embryonic stem cell marker SSEA-3, SSEA-4, TRA-1-60 and TRA-1-81 (Kisiel et al., 2012; Kang et al., 2012). In this study, 3th passage cAMSCs were identified by flow cytometry, the results showed that, cAMSCs had high expression CD29, CD49d and CD73, low expression CD34, CD45 and HLA-DR, consistent with the above results. The stem cell marker genes OCT4, SOX2 and NANOG are mainly expressed in embryonic stem cells, which play an important role in maintaining the pluripotent differentiation potential and self-renewal capacity of embryonic stem cells (Adewumi et al., 2007). In this study, we demonstrated that cAMSCs expressed OCT4, SOX2 and NANOG by RT-PCR. These results indicate that isolated the genes and proteins related to stem cells which are high purity cAMSCs.
In conclusion, this experiment isolated from the higher purity of the cAMSCs, expression of stem cell marker proteins and genes, and has multi-directional differentiation potential. This experiment provides a new source of stem cells for veterinary clinical practice and provides experimental basis for the study of canine regenerative medicine.
3.3 Differentiation potential of cAMSCs
In order to explore the differentiation ability of cAMSCs, in this study, osteogenic and adipogenic differentiation was performed. So far, there is no uniform method for the differentiation of cAMSCs into osteogenic. Many studies have shown that, dexamethasone, vitamin C and sodium glycerophosphate are necessary conditions for stem cells to differentiate into osteogenic in vitro. Dexamethasone on osteogenic differentiation of cAMSCs have both to promote the role of inhibition, mainly with the concentration, duration of action and cell types, low concentration of dexamethasone can promote matrix synthesis and calcification, thereby promoting the osteogenic differentiation of cAMSCs, when the concentration of 10 nmol / L, its role in promoting osteogenic differentiation of the most obvious. Sodium glycerophosphate can synergize with dexamethasone, can provide phosphorus ion for alkaline phosphatase, promote organic phosphorus into inorganic phosphorus, accelerate calcium deposition and promote bone formation and calcification, mesenchymal stem cells are necessary for mineralization deposition condition. Vitamin C can influence the hydroxylation of lysine and proline at the end of the collagen, which can increase the activity of alkaline phosphatase, accelerate calcium deposition and promote the formation of calcified nodules. Alkaline phosphatase is an enzyme secreted by osteoblasts, which is one of the specific markers of osteogenic differentiation. Alizarin red is an anionic dye that produces red complexes with calcium ions. So, alkaline phosphatase staining and alizarin red staining are often used for the detection of osteogenic differentiation (Girolamo et al., 2007). In this study, the osteogenic differentiation of cAMSCs was induced by osteogenic differentiation in vitro using 10 nmol / L dexamethasone, 10 mmol / L β-phosphoglycerol and 50 μmol / L vitamin C. After 28 days of osteogenic induction, the cells Accumulation growth, morphological changes were not obvious. By alkaline phosphatase staining, showing a large number of purple staining. By alizarin red staining, showing dyed dark red mineralized nodules. It was detected by RT-PCR, expression of osteoblast marker gene collagen type I alpha 1 after cAMSCs osteogenic differentiation. These results suggest that cAMSCs can differentiate into osteoblasts in vitro.
Dexamethasone, indomethacin, 3-isobutyl-1-methylxanthine and insulin are classic inducers of adipogenic differentiation. When the concentration of dexamethasone was 1 μmol / L, it could activate the glucocorticoid receptor on the surface of stem cells and promote its differentiation into adipocytes. Indomethacin activates the intracellular PPARγ pathway and increases the expression of intracellular adipogenic inducer cAMP. Insulin can bind to IGF-1 receptor, activate ERK1 / 2 signal pathway, thereby increasing the expression of PPARγ, so that the formation of fat cells increased. Oil Red O is a fat-soluble dye that can be specifically stained with triglycerides in adipose tissue cells (Aguiari et al., 2008). In this study, cAMSCs were in vitro adipogenic differentiation cultured in medium containing 1 μmol / L dexamethasone, 10 mg / L insulin, 0.5 mmol / L 3-isobutyl-1-methylxanthine and 100 μmol / L indomethacin. After 21 days of adipogenic diifferentiation, the cell morphology no longer showed long spindle shape, but tends to polygonal or oval, intracellular high-refractive small round drops, cell body increased. By oil red O staining, showing a large number of cells were stained into small red fat droplets. Lipoprotein lipase is an important factor in promoting adipocyte differentiation. In preadipocytes expresstion. It is one of the classic markers of adipogenic differentiation of stem cells. In this study, RT-PCR analysis revealed that adipocyte marker gene LPL was expressed in cAMSCs after adipogenic differentiation, indicating that the induction protocol used can successfully induce cAMSCs into adipocytes in vitro.
4 Conclusion
The cAMSCs were successfully isolated from the amnion tissue of the dog by enzyme digestion; cAMSCs expressed stromal cells marker protein Vimentin and stem cell marker protein SSEA-4; cAMSCs had high expression stem cell marker CD29, CD49d and CD73, low expression CD34, CD45 and HLA-DR; cAMSCs under in vitro specific inducing conditions, can be successfully differentiated into osteogenesis and adipogenesis.
Acknowledgements
First of all, thanks to my mentor Professor Cao Guifang, in my experiments to give me the help and support. Second, thanks to Inner Mongolia maternal and child health hospital Bai Liheng teacher. Finally, thanks to my partner to help my experiment. Fund: National "863" Program Supported Projects (No.2008AA101005)
Aguiari P., Leo S., Zavan B., Vindigni V., Rimessi A., Bianchi K., Franzin C., Cortivo R., Rossato M., Vettor R., Abatangelo G., Pozzan T., Pinton P., and Rizzuto R., 2008, High glucose induces adipogenic differentiation of muscle-derived stem cells[J], Proc Natl. Acad. Sci. U S A, 105(4): 1226-1231
https://doi.org/10.1073/pnas.0711402105
Bárcena A., Muench M.O., Kapidzic M., Gormley M., Goldfien G.A., Fisher S.J., 2011, Human placenta and chorion: potential additional sources of hematopoietic stem cells for transplantation[J], Transfusion, 51 Suppl 4
Chu C. R., Szczodry M., Bruno S., 2010, Animal models for cartilage regeneration and repair, Tissue Eng. Part B Rev., Feb, 16: 105–115
Coulombe P.A., Wong P., 2004, Cytoplasmic intermediate filaments revealed as dynamic and multipurpose scaffolds[J], Nat Cell Biol, 6(8): 699-706
https://doi.org/10.1038/ncb0804-699
Crovace A., Favia A., Lacitignola L., Di Comite M. S., Staffieri F., Francioso E., 2008, Use of autologous bone marrow mononuclear cells and cultured bone marrow stromal cells in dogs with orthopaedic lesions, Veterinary research communications, Sep., 32: S39–S44
Csaki C., Matis U., Mobasheri A., Ye H., and Shakibaei M., 2007, Chondrogenesis, osteogenesis and adipogenesis of canine mesenchymal stem cells: a biochemical, morphological and ultrastructural study, Histochemistry and cell biology, 128(6):507-520
https://doi.org/10.1007/s00418-007-0337-z
Csaki C., Matis U., Mobasheri A., Ye H., and Shakibaei M.,2007, Chondrogenesis, osteogenesis and adipogenesis of canine mesenchymal stem cells: a biochemical, morphological and ultrastructural study. Histochemistry and cell biology, 128(6), 507-520
https://doi.org/10.1007/s00418-007-0337-z
Diaz-Prado S., Muinos-Lopez E., Hermida-Gomez T., Rendal-Vazquez M.E., Fuentes-Boquete I., de Toro F. J., and Blanco F.J., 2010, Isolation and Characterization of Mesenchymal Stem Cells from Human Amniotic Membrane, Tissue Eng Part C Methods
Dominici M., Le Blanc K., Mueller I., Mueller I., Slaper-Cortenbach I., Marini F.C., Krause D.S., Deans R. J., Keating A., Prockop D. J. and Horwitz E. M., 2006, Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement[J], Cytotherapy, 8: 315–317
https://doi.org/10.1080/14653240600855905
Dominici M., Le Blanc K., Mueller I., Slaper-Cortenbach I., Marini F., Krause D., Deans R., Keating A., Prockop D.J. and Horwitz E., 2006, Minimal criteria for defining multipotent mesenchymal stromal cells, The International Society for Cellular Therapy position statement, Cytotherapy 8, 315
https://doi.org/10.1080/14653240600855905
Fortier L.A., 2005, Stem cells: classifications, controversies, and clinical applications [J], Vet Surg, 34(5): 415-423
https://doi.org/10.1111/j.1532-950X.2005.00063.x
Girolamo L., Sartori M.F., Albisetti W. and Brini A.T., 2007, Osteogenic differentiation of human adipose-derived stem cells: comparison of two different inductive media[J], J Tissue Eng. Regen Med, 1(2): 154-157
https://doi.org/10.1002/term.12
Guercio A., Di Marco P., Casella S., Cannella V., Russotto L., Purpari G., Bella S.Di., Production of canine mesenchymal stem cells from adipose tissue and their application in dogs with chronic osteoarthritis of the humeroradial joints [J], Cell Biol. Int., 2012, 36(2): 189–194
https://doi.org/10.1042/CBI20110304
Guercio A., Di Marco P., Casella S., Cannella V., Russotto L., Purpari G., Di Bella S., Piccione G., 2012, Production of canine mesenchymal stem cells from adipose tissue and their application in dogs with chronic osteoarthritis of the humeroradial joints, Cell Biol. Int., Feb., 36:189–194
Hak-Hyun R. Y. U., Byung-Jae K. A. N. G., Sung-Su P. A. R. K., Yongsun K. I. M., Heung-Myong W. O. O., KIM W. H., and KWEON O. K., 2012, Comparison of mesenchymal stem cells derived from fat, bone marrow, Wharton’s jelly, and umbilical cord blood for treating spinal cord injuries in dogs. Journal of Veterinary Medical Science, 74(12): 1617-1630
https://doi.org/10.1292/jvms.12-0065
Kang B.J., Ryu H.H., Park S.S., Koyama Y., Kikuchi M., Woo H.M., Kim W.H., and Kweon O.K., 2012, Comparing the osteogenic potential of canine mesenchymal stem cells derived from adipose tissues, bone marrow, umbilical cord blood, and Wharton’s jelly for treating bone defects[J], J Vet Sci, 13(3): 299–310
https://doi.org/10.4142/jvs.2012.13.3.299
Kisiel A.H., McDuffee L.A., Masaoud E., Bailey T.R., Gonzalez B.P.E., and Nino-Fong R., 2012, Isolation, characterization, and in vitro proliferation of canine mesenchymal stem cells derived from bone marrow, adipose tissue, muscle, and periosteum. American journal of veterinary research, 73(8): 1305-1317
https://doi.org/10.2460/ajvr.73.8.1305
Kortesidis A., Zannettino A., Isenmann S., Shi S., Lapidot T., Gronthos S., 2005, Stromal-derived factor-1 promotes the growth, survival, and development of human bone marrow stromal stem cells, Blood 105:3793–3801
https://doi.org/10.1182/blood-2004-11-4349
Li W., Wang G., Liang W., Kang K., Guo K., Zhang Y., 2014, Integrin β3 Is Required in Infection and Proliferation of Classical Swine Fever Virus, PLoS One, 9(10): e110911
https://doi.org/10.1371/journal.pone.0110911
Lim J.H., Byeon Y.E., Ryu H.H., Jeong Y.H., Lee Y.W., Kim W.H., Kang K.S., and Kweon O.K., 2007, Transplantation of canine umbilical cord blood-derived mesenchymal stem cells in experimentally induced spinal cord injured dogs[J], J Vet Sci, 8(3): 275-282
https://doi.org/10.4142/jvs.2007.8.3.275
Little C.B., Smith M.M., 2008, Animal models of osteoarthritis, Curr Rheumatol Rev., 4:1–8
https://doi.org/10.2174/157339708785133523
Murphy S. V, Rosli S., Acharya R., Mathias L., Lim R., Wallace E., Jenkin G., 2010, Amnion epithelial cell isolation and characterization for clinical use, Current Protocols Stem Cell Biol., Chapter 1: Unit 1E 6
Naggara O., Darsaut T.E., Salazkin I., Soulez G., Guilbert F., Roy D., Weill A., Gevry G., Raymond J., 2010, A new canine carotid artery bifurcation aneurysm model for the evaluation of neurovascular devices, American Journal of Neuroradiology, 31: 967–971
https://doi.org/10.3174/ajnr.A1929
Parolini O., Alviano F., Bagnara G. P., Bilic G., Bühring H.J., Evangelista M., ... and Miki T., 2008, Concise review: isolation and characterization of cells from human term placenta: outcome of the first international Workshop on Placenta Derived Stem Cells. Stem cells, 26(2): 300-311
https://doi.org/10.1634/stemcells.2007-0594
Schneider M.R., Wolf E., Braun J., Kolb H.J., Adler H., 2008, Canine embryo-derived stem cells and models for human diseases, Human molecular genetics, 17: R42–R47
Serikov V., Hounshell C., Larkin S., Green W., Ikeda H., Mark C. W., Kuypers F.A., 2009, Human term placenta as a source of hematopoietic cells, Exp. Biol. Med, 234: 813 – 823
https://doi.org/10.1093/hmg/ddn078
Takemitsu H., Zhao D., Yamamoto I., Harada Y., Michishita M., and Arai T., 2012, Comparison of bone marrow and adipose tissue-derived canine mesenchymal stem cells, BMC veterinary research, 8(1): 150
https://doi.org/10.1186/1746-6148-8-150
The International Stem Cell Initiative, 2007, Characterization of human embryonic stem cell lines by the international stem cell initiative[J], Nature Biotechnology, 25(7): 803-816
https://doi.org/10.1038/nbt1318
Tsuji H., Miyoshi S., Ikegami Y., Hida N., Asada H., Togashi I., ... and Mori T., 2010, Xenografted human amniotic membrane–derived mesenchymal stem cells are immunologically tolerated and transdifferentiated into cardiomyocytes, Circulation research, 106(10): 1613-1623
https://doi.org/10.1161/CIRCRESAHA.109.205260
Vieira N.M., Brandalise V., Zucconi E., Secco M., Strauss B.E., and Zatz M.,2010, Isolation, characterization, and differentiation potential of canine adipose-derived stem cells. Cell transplantation, 19(3): 279-289



. PDF(396KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Yukun Zhang
. Liheng Bai
. Yan Li
. Moning Liu
. Guifang Cao
. Dafu Zhi
Related articles
. Canine
. Amnion
. Mesenchymal stem cells
. Differentiation
. Veterinary cell therapy
Tools
. Email to a friend
. Post a comment